BAB I
PENDAHULUAN
1.1 Latar Belakang
Fotosintesis
merupakan cara atau proses tumbuhan dalam menghasilkan energi yang digunakan
untuk pertumbuhan dan perkembangannya. Setiap tumbuhan memiliki daur
fotosintesi yang berbeda-beda, sehingga dibedakan menjadi 3 jenis tumbuhan dan
daur fotosintesisnya. Jenis tumbuhan yang dibedakan yaitu tumbuhan C3, C4, dan
CAM.
Tumbuhan
Tumbuhan C4 dan CAM lebih adaptif di daerah panas dan kering dibandingkan
dengan tumbuhan C3. Namun tanaman C3 lebih adaptif pada kondisi kandungan CO2
atmosfer tinggi. Sebagian besar tanaman pertanian, seperti gandum, kentang,
kedelai, kacang-kacangan, dan kapas merupakan tanaman dari kelompok C3.
Tumbuhan
C4 jenis tumbuhan yang hidup di daerah panas seperti jagung, tebu,
rumput-rumputan. Tumbuhan memiliki kebiasaan saat siang hari mereka tidak
membuka stomatanya secara penuh untuk mengurangi kehilangan air melalui
evaporasi/transpirasi.
Tipe
crassulacean acid metabolism ( CAM) merupakan tipe tanaman yang mengambil CO2 pada
malam hari, dan mengunakannya untuk fotosistensis pada siang harinya. Tumbuhan
CAM yang dapat mudah ditemukan adalah nanas, kaktus, dan bunga lili.
Dari
pemaparan diatas, maka makalah ini akan membahas mengenai perbedaan tumbuhan C4
dan CAM serta daur fotosintesis tumbuhan tersebut.
1.2 Tujuan dan
Manfaat
Tujuan
Tujuan
dari makalah ini adalah mengetahui daur fotosintesis pada tumbuhan C4 dan
tumbuhan CAM.
Manfaat
Adapun
manfaat dari makalah ini sebagai berikut:
1.
Bagi penulis
Memehami materi dengan
baik mengenai daur fotosintesi tumbuhan C4 dan CAM.
2.
Bagi pembaca
Menjadi referansi tambahan mengenai
materi daur fotosintesis tumbuhan C4 dan CAM.
BAB II
PEMBAHASAN
Berdasarkan
tipe fotosintesis, tumbuhan dibagi ke dalam tiga kelompok besar, yaitu C3, C4,
dan CAM (crassulacean acid metabolism). Tumbuhan C4 dan CAM lebih adaptif di
daerah panas dan kering dibandingkan dengan tumbuhan C3. Namun tanaman C3 lebih
adaptif pada kondisi kandungan CO2 atmosfer tinggi. Sebagian besar tanaman
pertanian, seperti gandum, kentang, kedelai, kacang-kacangan, dan kapas merupakan
tanaman dari kelompok C3.
2.1 Definisi tanaman C3
·
Tumbuhan C3 merupakan
tumbuhan subtropis yang
menghasilkan glukosa dengan
pengolahan CO2 melalui siklus Calvin, yang melibatkan enzim Rubisco sebagai penambat CO2.
·
Tanaman C3 adalah spesies tanaman
yang menghasilkan 3 atom C dalam PGA sebagai produk utama awal pembakaran CO2(Sulisbury,
1995).
·
Tanaman C3 adalah tanaman yang
mempunyai lintasan atau siklus PCR (Photosynthetic Carbon Reduction) atau
sering disebut siklus calvin yang dapat menghasilkan asam organik yang
mengandung 3 atom C dan jaringan yang terlibat dalam proses fotosintesis adalah
jaringan mesofil. Lintasan itu dimulai dari pengikatan CO2 dengan
RBP dan RuBP(Sitompul, 1995).
·
Tanaman C3 adalah kelompok tumbuhan
yang menghasilkan senyawa phospho gliseric acid yang memiliki 3 atom C pada
proses fiksasi CO2 oleh ribolusa diphosphat(Budiarti, 2008).
·
C3 tanaman melibatkan langsung fiksasi karbon CO 2. Artinya, langkah
awal melibatkan
CO2 yang terikat ribulosa bifosfat untuk menghasilkan dua molekul dari
tiga-karbon senyawa (yaitu
3-phosphogylycerate). Kunci
Pada
tanaman C3, enzim yang menyatukan CO2 dengan RuBP (RuBP merupakan substrat
untuk pembentukan karbohidrat dalam proses fotosintesis) dalam proses awal
assimilasi, juga dapat mengikat O2 pada saat yang bersamaan untuk proses
fotorespirasi ( fotorespirasi adalah respirasi,proses pembongkaran karbohidrat
untuk menghasilkan energi dan hasil samping, yang terjadi pada siang hari) .
Jika konsentrasi CO2 di atmosfir ditingkatkan, hasil dari kompetisi antara CO2
dan O2 akan lebih menguntungkan CO2, sehingga fotorespirasi terhambat dan
assimilasi akan bertambah besar.
Tumbuhan
C3 tumbuh dengan karbon fiksasi C3 biasanya tumbuh dengan baik di area dimana
intensitas sinar matahari cenderung sedang, temperature sedang dan dengan
konsentrasi CO2 sekitar 200 ppm atau lebih tinggi, dan juga
dengan air tanah yang berlimpah. Tumbuhan C3 harus berada dalam area dengan konsentrasi
gas karbondioksida yang tinggi sebab Rubisco sering menyertakan molekul oksigen
ke dalam Rubp sebagai pengganti molekul karbondioksida. Konsentrasi gas
karbondioksida yang tinggi menurunkan kesempatan Rubisco untuk menyertakan
molekul oksigen. Karena bila ada molekul oksigen maka Rubp akan terpecah
menjadi molekul 3-karbon yang tinggal dalam siklus Calvin, dan 2 molekul
glikolat akan dioksidasi dengan adanya oksigen, menjadi karbondioksida yang
akan menghabiskan energi.
Pada
tumbuhan C3, CO2 hanya difiksasi RuBP oleh karboksilase RuBP. Karboksilase RuBP
hanya bekerja apabila CO2 jumlahnya berlimpah
Contoh tanaman C3 antara lain : kedelai, kacang tanah, kentang, dll.
Contoh tanaman C3 antara lain : kedelai, kacang tanah, kentang, dll.
Fiksasi Karbondioksida
Melvin Calvin bersama
beberapa peneliti pada universitas calivornia berhasil mengidentivikasi produk
awal dari fiksasi CO2. Produk awal tersebut adalah asam
3-fosfogliserat atau sering disebut PGA, karena PGA tersusun dari 3 atom
karbon.
Hasil penelitian menunjukkan bahwa tidak ada senyawa dengan 2
atom C yang terakumulasi. Senyawa yang terakumulasi adalah senyawa dengan 5
atom C yakni Ribulosa – 1.5 – bisfosfat (RUBP). Reaksi antara CO2 dengan
RUBP dipacu oleh enzim ribulosa bisfosfat karboklsilase (RUBISCO).
Rubisco adalah enzim raksasa
yang berperan sangat penting dalam reaksi gelap fotosintesis tumbuhan.
Enzim inilah yang menggabungkan molekul
ribulosa-1,5-bisfosfat (RuBP,
kadang-kadang disebut RuDP) yang memiliki tiga atom C dengan karbondioksida menjadi
atom dengan enam C, untuk kemudian diproses lebih lanjut menjadi glukosa,
molekul penyimpan energi aktif utama pada tumbuhan.
Siklus Calvin
Siklus Calvin disebut
juga Reaksi gelap yang merupakan
reaksi lanjutan dari reaksi terang dalam fotosintesis.
Reaksi gelap adalah reaksi pembentukan gula dari CO2 yang terjadi di stroma. Reaksi
ini tidak membutuhkan cahaya. Reaksi terjadi pada bagian kloroplas yang
disebut stroma.
Tempat terjadinya Reaksi gelap
Bahan reaksi gelap
adalah ATP dan NADPH, yang dihasilkan dari reaksi terang, dan CO2,
yang berasal dari udara bebas. Dari reaksi gelap ini, dihasilkan glukosa (C6H12O6),
yang sangat diperlukan bagi reaksi katabolisme. Reaksi ini ditemukan oleh Melvin
Calvin dan Andrew Benson, karena itu reaksi
gelap disebut juga reaksi Calvin-Benson.
Secara umum, reaksi
gelap dapat dibagi menjadi tiga tahapan (fase), yaitu fiksasi, reduksi, dan
regenerasi. Reaksi gelap dimulai dengan pengikatan atau fiksasi 6 molekul CO2
ke 6 molekuk gula 5 karbon yaitu ribulosa 1,5 bifosfat, dikatalisis oleh enzim
ribulosa bifosfat karboksilase/oksigenase(rubisco) yang kemudian membentuk 6
molekul gula 6 karbon. Molekul 6 karbon ini tidak stabil maka pecah menjadi 12
molekul 3 karbon yaitu 3 fosfogliserat. 3 fosfogliserat kemudian difosforilasi
oleh 12 ATP membentuk 1,3 bifosfogliserat. 1,3 bifosfogliserat difosforilasi
lagi oleh 12 NADPH membentuk 12 molekul gliseradehida 3 fosfat/PGAL. 2 PGAL
digunakan untuk membentuk 1 molekul glukosa atau jenis gula lainnya, sedangkan
10 molekul lainnya difosforilasi oleh 6 ATP untuk kembali membentuk 6 molekul
Ribulosa 1,5 bifosfat. Proses pengikatan CO2 ke RuBP disebut fiksasi,
proses pemecahan molekul 6 karbon menjadi molekul 3 karbon disebut reduksi dan
proses pembentukan kembali RuBP dari PGAL disebut regenerasi.
Fotosintesis ini
disebut mekanisme C3, karena molekul yang pertama kali terbentuk setelah
fiksasi karbon adalah molekul berkarbon 3, 3-fosfogliserat. Kebanyakan tumbuhan
yang menggunakan fotosintesis C3 disebut tumbuhan C3.
Padi, gandum, dan kedelai merupakan
contoh-contoh tumbuhan C3 yang penting dalam pertanian. Kondisi
lingkungan yang mendorong fotorespirasi ialah hari yang panas, kering, dan
terik-kondisi yang menyebabkan stomata tertutup. Kondisi ini menyebabkan CO2
tidak bisa masuk dan O2 tidak bisa keluar sehingga terjadi fotorespirasi.
2.2 Definisi
tanaman C4
·
Tumbuhan C4 adalah
tumbuhan tropis yang
melibatkan dua enzim di dalam pengolahan CO2 menjadi glukosa
yaitu Enzim phosphophenol pyruvat carboxilase (PEPco) adalah enzim yang akan
mengikat CO2 dari udara dan kemudian akan menjadi oksaloasetat yang akan diubah menjadi
malat.
·
Tanaman C4 adalah tanaman yang
menghasilkan asam 4 karbon sebagai produk utama penambahan CO2(Salisburry,
1998).
·
Tanaman C4 adalah kelompok tumbuhan
yang melakukan persiapan reaksi gelap fotosintesis melalui jalur 4 karbon / 4C
(jalur hatch- slack) sebelum memasuki siklus calvin, untuk meminimalkan
keperluan fotorespirasi
(Budiarti,
2008).
·
Tanaman C4 adalah tanaman dengan
hasil pertama dalam fotosintesis di mesofil berupa suatu molekul dengan 4 atom
C(Gardner, 1991).
Tumbuhan C4 dan CAM lebih adaptif di
daerah panas dan kering. Pada tanaman C4, CO2 diikat oleh PEP (enzym pengikat
CO2 pada tanaman C4) yang tidak dapat mengikat O2 sehingga tidak terjadi
kompetisi antara CO2 dan O2. Lokasi terjadinya assosiasi awal ini adalah di
sel-sel mesofil (sekelompok sel-sel yang mempunyai klorofil yang terletak di
bawah sel-sel epidermis daun). CO2 yang sudah terikat oleh PEP kemudian
ditransfer ke sel-sel “bundle sheath” (sekelompok sel-sel di sekitar xylem dan
phloem) dimana kemudian pengikatan dengan RuBP terjadi. Karena tingginya
konsentasi CO2 pada sel-sel bundle sheath ini, maka O2 tidak mendapat
kesempatan untuk bereaksi dengan RuBP, sehingga fotorespirasi sangat kecil and
G sangat rendah, PEP mempunyai daya ikat yang tinggi terhadap CO2, sehingga
reaksi fotosintesis terhadap CO2 di bawah 100 m mol m-2 s-1 sangat tinggi. ,
laju assimilasi tanaman C4 hanya bertambah sedikit dengan meningkatnyaCO2.
Sehingga, dengan meningkatnya CO2 di atmosfir, tanaman C3 akan lebih beruntung
dari tanaman C4 dalam hal pemanfaatan CO2 yang berlebihan. Contoh tanaman C4
adalah jagung, sorgum dan tebu.
Tetapi pada sintesis C4,enzim
karboksilase PEP memfiksasi CO2 pada akseptor karbon lain yaitu PEP.
Karboksilase PEP memiliki daya ikat yang lebih tinggi terhadap CO2 daripada
karboksilase RuBP. Oleh karena itu,tingkat CO2 menjadi sangat rendah pada
tumbuhan C4,jauh lebih rendah daripada konsentrasi udara normal dan CO2 masih
dapat terfiksasi ke PEP oleh enzim karboksilase PEP. Sistem perangkap C4 bekerja
pada konsentrasi CO2 yang jauh lebih rendah.
Tumbuhan C4 dinamakan demikian karena
tumbuhan itu mendahului siklus Calvin yang menghasilkan asam berkarbon -4
sebagai hasil pertama fiksasi CO2 dan yang memfiksasi CO2 menjadi
APG di sebut spesies C3, sebagian spesies C4 adalah
monokotil (tebu, jagung, dll).
Reaksi dimana CO2 dikonfersi
menjadi asam malat atau asam aspartat adalah melalui penggabungannya dengan
fosfoeolpiruvat (PEP) untuk membentuk oksaloasetat dan Pi. Enzim
PEP-karboksilase ditemukan pada setiap sel tumbuhan yang hidup dan enzim ini
yang berperan dalam memacu fiksasi CO2 pada tumbuhan C4.
Enzim PEP-karboksilase terkandung dalam jumlah yang banyak pada daun tumbuhan C4,
pada daun tumbuhan C-3 dan pada akar, buah-buah dan sel – sel tanpa klorofil
lainnya ditemukan suqatu isozim dari PEP-karboksilase.
Reaksi untuk mengkonversi oksaloasetat menjadi malat
dirangsang oleh enzim malat dehidrogenase dengan kebutuhan elektronnya
disediakan oleh NHDPH. Oksaleasetat harus masuk kedalam kloroplas untuk direduksi
menjadi malat.
Pembentukkan aspartat dari malat terjadi didalam
sitosol dan membutuhkan asam amino lain sebagai sumber gugus aminonya. Proses
ini disebut transaminasi.
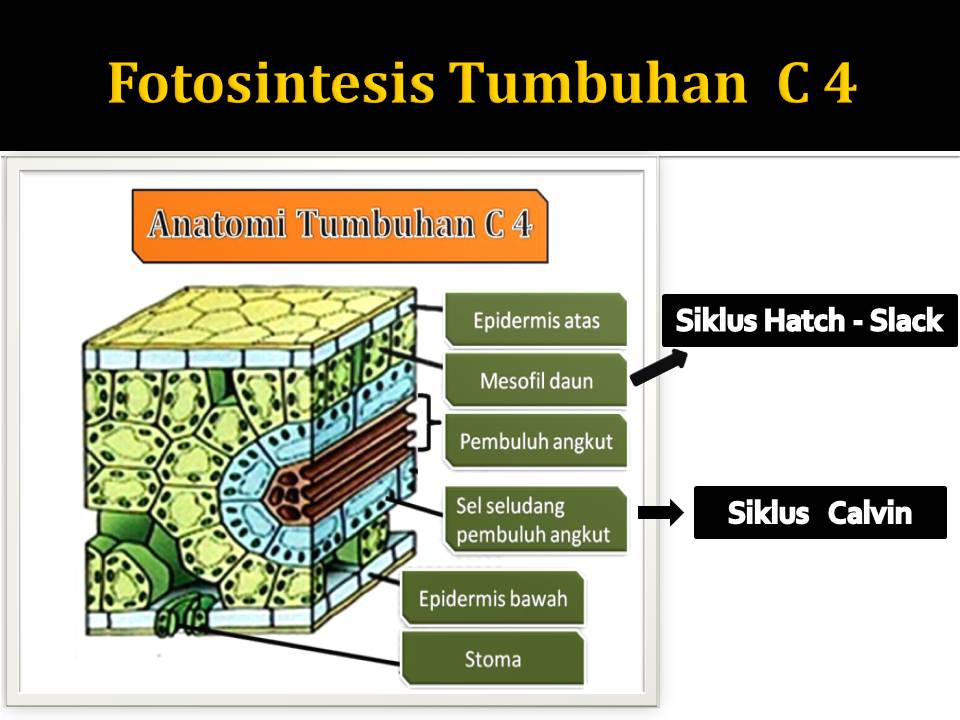
Pada tumbuhan C-4 terdapat pembagian tugas antara 2
jenis sel fotosintetik, yakni:
- sel
mesofil
- sel-sel
bundle sheath/ sel seludang-berkas pembuluh.
Sel seludang berkas pembuluh disusun menjadi kemasan
yang sangat padat disekitar berkas pembuluh. Diantara seludang-berkas pembuluh
dan permukaan daun terdapat sel mesofil yang tersusun agak longgar. Siklus
calvin didahului oleh masuknya CO2 ke dalam senyawa organik
dalam mesofil.
Langkah pertama ialah penambahan CO2 pada
fosfoenolpirufat (PEP) untuk membentuk produk berkarbon empat yaitu
oksaloasetat, Enzim PEP karboksilase menambahkan CO2 pada PEP.
Karbondioksida difiksasi dalam sel mesofil oleh enzim PEP karboksilase. Senyawa
berkarbon-empat-malat, dalam hal ini menyalurkan atom CO2 kedalam
sel seludang-berkas pembuluh, melalui plasmodesmata. Dalam sel seludang –berkas
pembuluh, senyawa berkarbon empat melepaskan CO2 yang
diasimilasi ulang kedalam materi organic oleh robisco dan siklus Calvin. Dengan
cara ini, fotosintesis C4 meminimumkan fotorespirasi dan
meningkatkan produksi gula. Adaptasi ini sangat bermanfaat dalam daerah panas
dengan cahaya matahari yang banyak, dan dilingkungan seperti inilah tumbuhan C4 sering
muncul dan tumbuh subur.
2.3 Definisi
tanaman CAM
·
Tanaman CAM adalah tanaman yang
dapat berubah seperti tanaman C3 pada saat pagi hari (suhu rendah) dan dapat
berubah seperti tanaman C4 pada siang hari dan malam hari(Gardner, 1991).
·
Tanaman CAM adalah tanaman yang
membuka pada malam hari dan menutup pada siang hari, memiliki laju fotosintesis
yang rendah bila dibandingkan dengan tanaman C3 dan C4(Lakitan, 1995).
Tanaman CAM , pada kelompok ini
penambatan CO2 seperti pada tanaman C4, tetapi dilakukan pada malam hari dan
dibentuk senyawa dengan gugus 4-C. Pada hari berikutnya ( siang hari ) pada
saat stomata dalam keadaan tertutup terjadi dekarboksilase senyawa C4 tersebut
dan penambatan kembali CO2 melalui kegiatan Rudp karboksilase. Jadi tanaman CAM
mempunyai beberapa persamaan dengan kelompok C4 yaitu dengan adanya dua tingkat
sistem penambatan CO2.
Pada C4 terdapat pemisahan ruang sedangkan pada CAM
pemisahannya bersifat sementara. Termasuk golongan CAM adalah Crassulaceae, Cactaceae, Bromeliaceae,
Liliaceae, Agaveceae, Ananas comosus, dan Oncidium lanceanum.
Beberapa tanaman CAM dapat beralih ke jalur C3 bila keadaan lingkungan lebih baik.
Beberapa tanaman CAM dapat beralih ke jalur C3 bila keadaan lingkungan lebih baik.
Beberapa spesies tumbuhan mempunyai
sifat yang berbeda dengan kebanyakan tumbuhan lainnya, yakni tumbuhan ini
membuka stomatanya pada malam hari dan menutupnya pada siang hari. Kelompok
tumbuhan ini umumnya adalah tumbuhan jenis sukulen yang tumbuh di daerah
kering. Dengan menutup stomata pada siang hari membantu tumbuhan ini menghemat
air, dapat mengurangi laju transpirasinya, sehingga lebih mampu beradaptasi
pada daerah kering tersebut.
Selama malam hari, ketika stomata
tumbuhan itu terbuka, tumbuhan ini mengambil CO2 dan memasukkannya
kedalam berbagai asam organik. Cara fiksasi karbon ini disebut metabolisme
asam krasulase, atau crassulacean acid metabolism
(CAM). Dinamakan demikian karena metabolisme ini pertama kali
diteliti pada tumbuhan dari famili crassulaceae. Jalur CAM serupa dengan jalur
C4 dalam hal karbon dioksida terlebih dahulu dimasukkan kedalam
senyawa organik intermediet sebelum karbondioksida ini memasuki siklus Calvin.
Perbedaannya ialah bahwa pada tumbuhan C4, kedua langkah ini terjadi
pada ruang yang terpisah. Langkah ini terpisahkan pada dua jenis sel. Pada
tumbuhan CAM, kedua langkah dipisahkan untuk sementara. Fiksasi karbon terjadi
pada malam hari, dan siklus calvin berlangsung selama siang hari.
2.4 Karakteristik
tanaman CAM
Tanaman CAM adalah tumbuhan sukulen yang pada umumnya
tidak memiliki lapisan sel palisade yang teratur. Sel daun dan ranting
merupakan sel mesofil bunga karang. Terdapat sel bundle sheath tetapi sel
tersebut tidak banyak berbeda dengan sel mesofil. Pada CAM, pembentukan asam
malat pada malam hari, dibarengi dengan penguraian gula, pati, atau polimer
glukosa yang mirip dengan pati.
Tanaman CAM (Crassulation Acid Metabolism Plants) pada
dasarnya adalah tanaman sukulen yaitu tanaman yang berdaun atau berbatang tebal
yang bertranspirasi rendah. Dalam kondisi kering, stomata pada malam hari akan
terbuka untuk mengabsorbsi CO2dan menutup pada siang hari untuk mengurangi
transpirasi. Fiksasi CO2 tanaman CAM sama seperti tanaman C4,
hanya saja terjadinya pada malam hari dan energi yang dibutuhkan diperoleh dari
glikolisis. Namun dalam kondisi cukup lemah, banyak spesies CAM merubah
fungsistomata dan karboksilasi seperti tanaman C3. Tanaman CAM juga mempunyai
metode fisiologis untuk mereduksi kehilangan air dan menghindari kekeringan.
(Salisburry, 1998).
sintesis C4
Pada jenis tumbuhan yang hidup di daerah panas seperti
jagung, tebu, rumput-rumputan, memiliki kebiasaan saat siang hari mereka tidak
membuka stomatanya secara penuh untuk mengurangi kehilangan air melalui
evaporasi/transpirasi. Ini berakibat terjadinya penurunan jumlah CO2 yang masuk
ke stomata. Logikanya hal ini menghambat laju fotosintesis. Ternyata para
tumbuhan ini telah mengembangkan cara yang cerdas untuk menjaga agar laju
fotosintesis tetap normal meskipun stomata tidak membuka penuh.
Perbedaannya ada pada mekanisme fiksasi CO2. Pada
tumbuhan C-4 karbondioksida pertamakali akan diikat oleh senyawa yang disebut
PEP (phosphoenolphyruvate / fosfoenolpiruvat) dengan bantuan enzim PEP
karboksilase dan membentuk oksaloasetat, suatu senyawa 4-C. Itu sebabnya
kelompok tumbuhan ini disebut tumbuhan C-4 atau C-4 pathway. PEP dibentuk dari
piruvat dengan bantuan enzim piruvat-fosfat dikinase. Berbeda dengan rubisco,
PEP sangat lemah berikatan dengan O2. Ini berarti bisa menekan terjadinya
fotorespirasi sekaligus mampu menangkap lebih banyak CO2 sehingga bisa meningkatkan
laju produksi glukosa(Lehninger, 1982).
Pengikatan CO2 oleh PEP tersebut berlangsung di
sel-sel mesofil (daging daun). Oksaloasetat yang terbentuk kemudian akan
direduksi karena menerima H+ dari NADH dan berubah menjadi malat, kemudian
ditransfer menuju ke sel seludang pembuluh (bundle sheath cells) melalui
plasmodesmata. Sel-sel seludang pembuluh adalah kelompok sel yang mengelilingi
jaringan pengangkut xilem dan floem. Lihat gambar.
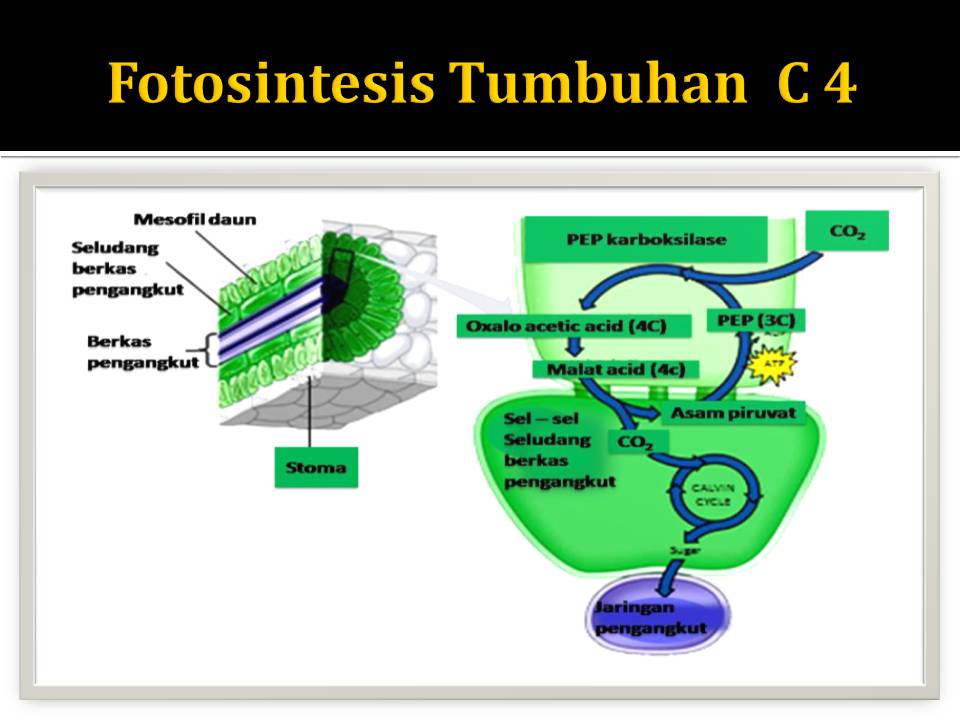

Di dalam
sel-sel seludang pembuluh malat akan dipecah kembali menjadi CO2 yang langsung
memasuki siklus Calvin-Benson, dan piruvat dikembalikan lagi ke sel-sel
mesofil. Hasil dari siklus Calvin-Benson adalah molekul glukosa yang kemudian
ditranspor melalui pembuluh floem.
Dari uraian di atas kita tahu bahwa fiksasi CO2 pada
tumbuhan C-4 berlangsung dalam dua langkah. Pertama CO2 diikat oleh PEP menjadi
oksaloasetat dan berlangsung di sel-sel mesofil. Kedua CO2 diikat oleh rubisco
menjadi APG di sel seludang pembuluh. Ini menyebabkan energi yang digunakan
untuk fiksasi CO2 lebih besar, memerlukan 30 molekul ATP untuk pembentukan satu
molekul glukosa. Sedangkan pada tumbuhan C-3 hanya memerlukan 18 molekul ATP.
Namun demikian besarnya kebutuhan ATP untuk fiksasi CO2 pada tumbuhan C-4
sebanding dengan besarnya hasil produksi glukosa karena dengan cara tersebut
mampu menekan terjadinya fotorespirasi yang menyebabkan pengurangan pembentukan
glukosa. Itu sebabnya kelompok tumbuhan C-4 dikenal efektif dalam fotosintesis.
Sintesis CAM
Tumbuhan
lain yang tergolong sukulen (penyimpan air) misalnya kaktus dan nanas memiliki
adaptasi fotosintesis yang berbeda lagi. Tidak seperti tumbuhan umumnya,
kelompok tumbuhan ini membuka stomata pada malam hari dan menutup pada siang
hari. Stomata yang menutup pada siang hari membuat tumbuhan mampu menekan
penguapan sehingga menghemat air, tetapi mencegah masuknya CO2.

Saat
stomata terbuka pada malam hari, CO2 di sitoplasma sel-sel mesofil akan diikat
oleh PEP dengan bantuan enzim PEP karboksilase sehingga terbentuk oksaloasetat
kemudian diubah menjadi malat (persis seperti tumbuhan C-4). Selanjutnya malat
yang terbentuk disimpan dalam vakuola sel mesofil hingga pagi hari. Pada siang
hari saat reaksi terang menyediakan ATP dan NADPH untuk siklus Calvin-Benson,
malat dipecah lagi menjadi CO2 dan piruvat. CO2 masuk ke siklus Calvin-Benson
di stroma kloroplas, sedangkan piruvat akan digunakan untuk membentuk kembali
PEP.

Model
metabolisme ini disebut Crassulacean Acid Metabolism (CAM) karena
pertamakali diketahui terjadi pada kelompok tumbuhan famili Crassulaceae.
Jadi maksud penamaannya berarti: metabolisme asam pada tumbuhan
Crassulaceae dan bukan metabolisme asam Crassulaceae(Lakitan,
2007).
BAB III
SIMPULAN
1.
Tumbuhan C4 adalah tumbuhan tropis yang melibatkan dua enzim di
dalam pengolahan CO2 menjadi glukosa yaitu Enzim phosphophenol
pyruvat carboxilase (PEPco) adalah enzim yang akan mengikat CO2 dari
udara dan kemudian akan menjadi oksaloasetat yang akan diubah menjadi
malat.
2.
Tanaman CAM adalah tanaman yang
membuka pada malam hari dan menutup pada siang hari, memiliki laju fotosintesis
yang rendah bila dibandingkan dengan tanaman C3 dan C4.
DAFTAR PUSTAKA
Budiarti. 2008. Pengantar
Fisiologi Tumbuhan. Gramedia, Jakarta
Gardner. 1991. Fisiologi
Tanaman Budidaya. UI Press. Jakarta
Lakitan, 1995. Dasar-dasar
Fisiologi Tumbuhan. Rajawali Grafindo, Jakarta
Lakitan,
Benyamin. 2007. Dasar-Dasar Fisiologi Tumbuhan. PT.Raja Grafindo Persada.
Jakarta
Lehninger, Albert . L.
1982. Dasar-Dasar Biokimia. Penerbit Erlangga
Salisbury,
Frank. B dan C.W. Ross. 1995. Fisiologi Tumbuhan. Penerbit ITB.Bandung
Sitompul,
SM. 1995. Fisiologi Tanaman Tropis. Universitas Mataram. Lombok.



0 komentar:
Posting Komentar